Autres cours d'SVT
La cellule d'ADN et l'Unité du vivant
Diversité et parenté des
vertébrés.
Le coeur
Chapitre 3 : Diversité et parenté des Vertébrés.
I) Organisation des Vertébrés.
II) Origine du plan d'organisation.
A) Développement de l'embryon.
1)De la cellule œuf (union des deux gamètes —> 1+1=1) à la neurula.
2) Développement embryonnaire après la neurula.B) L'apparition des axes de polarité.
1) Document "ovocyte d'oursin" avant la fécondation.
2) Point d'entrée du spermatozoïdeC. Mise en place des différentes parties du corps.
1) p.241 : expérience des tissus transplanté.
2) expérience des ailes de poulet.
III) Contrôle génétique du plan d'organisation
I) Organisation des Vertébrés.
Diversité = nombre d'espèces animales et végétales.
Chacun est unique.
Parenté = évolution depuis origine.
A) Étude Morphologique
(voir feuille)
Les trois axes de polarité définissent la parité chez les vertébrés.
B) Étude anatomique.
C'est l'ensemble des caractéristiques morphologiques, anatomiques et physiologiques présents chez tous les vertébrés => plan d'organisation.
Voir TP.1 : Organisation des Vertébrés.
II) Origine du plan d'organisation.
A) Développement de l'embryon.
1)De la cellule œuf (union des deux gamètes —> 1+1=1) à la neurula.
L'ovule comme la cellule-oeuf sont nichés en réserve (le vitellus) qui servira à la construction des autres cellules puis de l'embryon.
Étapes du développement de la cellule oeuf jusqu'à la neurula:
- la segmentation (division binaire)
- la morula (petite sphère constituée de quelque millier de cellules)
- la blastula qu'on reconnaît grâce à une cavité
centrale, le blastocoele. Certaines cellules vont commencer `a migrer
et se regrouper à un pôle.
- la gastrula (3 feuillets représentés par 3 couleurs
différentes) :
- le bleu : l'ectoderme (à l'extérieur) ce qui donnera de la peau et le système nerveux.
- le rouge : le mésoderme (au milieu) donnera les muscles ainsi que le système reproductif.
- le jaune : l'éctoderme (à l'intérieur) deviendra
le système digestif et l'appareil respiratoire.
- Neurula : a partir de ce stade, commence la différenciation des cellules.
2) Développement embryonnaire après la neurula.
Les 5 classes de vertébrés (les Amphibiens, les mammifères, les poissons, les oiseaux et les reptiles) présentent dès l'origine les 3 axes de polarité, les mêmes caractéristiques morphologiques et donc ont tous le même plan d'organisation => parenté.
B) L'apparition des axes de polarité.
Ces axes ne semblent pas apparent lors des premiers stades embryonnaires. Qu'en est-il?
1. Document "ovocyte d'oursin" avant la fécondation.
Étape d'étude :
- matériel —> Ce que j'en fais.
expérience—> comment je la fais. - résultat—> ce que je vois
- conclusion —> ce que j'en pense.
L'ovocyte est un futur ovule. Il n'est pas entièrement fini, il lui manque des réserves.
Rq : L'ovule est de plusieurs couleurs (dégradé du noir au blanc).
- 2 ovocytes d'oursins (2 noyaux) indispensables pour l'information génétique —> 2 moitiés d'ovocyte nucléos pour être fécondés.
- La moitié supérieur donne un organisme rond alors que l'inférieur donne une larve mais aucune ne va donner un oursin entier.
- Les axes de polarité apparaissent des l'ovocyte. La partie supérieure donne la forme alors que l'inférieure les axes.
Les 2 parties d'un ovocyte sont déjà spécifiée pour donner les différents axes de polarité:
- la partie supérieur détermine la morphologie (la forme)
- la partie inférieur détermine les axe antéro-postérieur et latéral.
2. Point d'entrée du spermatozoïde
Un ovule est caractérisé par la présence de deux pôles :
- pôle animal
- pôle végétatif.
Dès l'entrée du spermatozoïde (près du pôle animal), apparaît un croissant gris du côté du pôle végétatif. Cette couleur grise vient de la dépigmentation de cette zone.
Ce croissant se déplace.
Dans l'expérience, on empêche se déplacement.
—> on obtient un têtard difforme.
Les axes de polarité sont donc aussi défini par le point d'entrée du spermatozoïde. Mais celui-ci doit rentrer dans le pôle animal de l'ovule.
Document 3. Les étapes précoces du développement embryonnaire.
- On voit les premiers stades de la segmentation de la cellule-oeuf. On sépare les deux moitiés d'un embryon de xénope au stade 4 cellule selon un axe "pôle animale pôle végétatif", une moitié contenant le croissant gris.
- La moitié contenant le croissant gris donne un embryon dosalisé mais sans tube digestif alors que l'autre moitié (sans le croissant gris) donne un embryon ventralisé mais sans tube nerveux.
- Les. Les axes de polarité sont donc déjà déterminéau stade 4 cellules. Le croissant est donc responsable de l'apparition des axes de polarité. C'est donc la répartition des réserves du cytoplasme qui va déterminerl'apparition des axes.
Étude du document 4
- On a deux embryons dont un témoin au stade 8 cellules. Sur l'un d'eux on effectue une rotation de 180° des quatre cellules du pole animal.
- On constate que l'embryon témoin se développe au stade
neurula avec une polarité originale, tandis que le 2ème
embryon au même stade à développé une polarité
inverse.
(c'est un peu comme si on avait la colonne vertébrale sur le ventre...) - On peut donc en déduire que l'axe dorso-ventral est déterminée par le pôle animal.
C. Mise en place des différentes parties du corps.
1) p.241 : expérience des tissus transplanté.
Lors de la première expérience le greffon est prélevésur une jeune gastrula et greffer dans une autre :
- lorsque le tissu transplanté (prédestiner a donner de la peau = éctoderme) est greffé dans la partie qui donnera du mésoderme, le greffon donnera du mésoderme
- de même quand ce même tissu est transplanté dans la région de la gasrula qui deviendra du cerveau, le greffon donnera du cerveau (mésoderme.
On peut donc déduire qu'au stade de la jeune neurula les cellules ne sont pas encore spécialisée, où que toute ces cellules possèdent le même génome qui ne s'exprime pas.
Lors de la deuxième expérience, on transplante le même greffon mais sur une gastrula âgée :
- lorsque le tissu transplanté (prédestiner a donner de la peau = éctoderme) est greffé dans la partie qui donnera du mésoderme, le greffon donnera non pas du mésoderme mais de l'éctoderme.
- de même quand le tissu transplanté (prédestiner a donner de la peau = éctoderme) est greffé dans la partie qui donnera du mésoderme, le greffon donnera ne donnera pas du mésoderme, mais de l'éctoderme.
Les cellules du greffon sont donc déjà déterminée et ont acquis leur spécification.
Conclusion des expériences : L'origine des différents organes et parties du corps sont donc mis en place dès le stade gastrula, d'où le passage à la morula.
2) expérience des ailes de poulet.
Voir feuille.
La présence de la crête sur le bourgeon est très importante, puisque l'ablation de cette crête provoque l'arrêt du développement du membre. L'aile de poulet se construit à partir du bourgeon, mais celui-ci ne réagit qu'en présence de la crête.
Quelle est la relation entre la crête le bourgeon?
Le FGF est une substance chimique capable de provoquer la croissance. Il y a une information qui passe de la crête au bourgeon; cette relation est hormonale.
Il existe donc des signaux chimiques émis et compris entre les cellules qui permettent la mise en place des différents organes.
III) Contrôle génétique du plan d'organisation
A) Importance de certains gènes.
document 1 sur feuille.
Chaque mutation est due à la mutation d'un seul gène particulier (ici chez la drosophile les gènes bicoïd, nanos et torso)
A chaque gène correspond le développement d'une partie précise de l'animal.
Chaque partie du corps se développe grâce la présence d'un gène bien particulier appelé, gène de développement ou gène homéotique.
document A sur la même feuille.
A chaque gène correspond la fabrication d'une protéine précise.
- la concentration d'une protéine de développement varie de la tête à la queue.
- le développement d'une partie du corps est soumis à une certaine quantité de protéines
Ici, chez le mutant, la quantité de protéines de développement est très basse et constante de la tête à la queue.
Conclusion : le développement d'un organisme s'exprime par les gènes qui s'expriment par l'intermédiaire des protéines. Le développement d'une partie du corps est relatif à la concentration des protéines.
document B toujours sur la même feuille.
On injecte les protéines de la tête d'un individu normal dans la partie de la tête d'un embryon muté et l'on obtient une larve avec une tête.
Conclusion : C'est effectivement la quantité de la protéine qui est responsable pour le développement d'une partie du corps.
B) Expérience de transgènese.
voir livre p.243.
La construction d'un organe demande l'intervention d'un gène principal. C'est le gène-architecte. Ce gène peut interagir avec beaucoup d'autres gènes qui ne s'expriment pas tant que le gène maître reste inactif.
Document 1 p.243
- On utilise le gène "Small eye"(qui sert à
fabriquer de l'oeil) d'une souris.
On utilise le gène "Eyeless" d'une drosophile (qui lui aussi sert à crée de l'oeil) - On active le gène "Eyeless" dans une cellule où
il est normalement inactif
Résultat, on a un oeil n'importe où.
On conclu que le gène "Eyeless" est architecte et qu'il permet l'organisation de l'oeil. - On greffe un gène "Small Eye" d'une souris dans une
drosophile.
Hypothèse : Sur la drosophile apparaîtra des yeux de souris
Résultat : l'oeil est normal, celui d'une drosophile
On conclu que le gène "Small eye" de la souris et le gène "Eyeless" de la drosophile sont homologues.
Que ce passe-t-il au niveau de l'ADN?
La gènètique du développement est une usine, où les maîtres sont les gènes architectes, les contre-maîtres, les gènes et les ouvries les protèines.
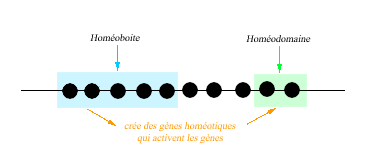
Document 2 p.245.
- Ensemble des gènes homéotiques répartis sur un
chromosome chez la drosophile.
Ensemble des gènes homéotiques répartis sur plusieurs chromosomes chez une souris - ressemblances :
- ordre
- 1er gène est celui pour la tête
Conclusion :
On constate que tout gènes homéotique de différentes
espèces possèdent des homéoboites identiques qui
fabriquent des homéodomaines identiques qui vont agir sur d'autre
gènes de la même façon, c'est à dire en activant
des gènes architectes.
2002, Jel